At first glance, evolutionary biology and astrophysics seem to have very little in common. However, they are united in that they suffer from similar fundamental predicament: The difficulty of observation and experiment. These difficulties manifest in that the observed phenomena have inconvenient timescales, cannot be fully repeated in a lab experiment, and may only have one realization. In the following, we will look at each of these separate problems in more detail.
Partly due to these issues, evolutionary biology, and in particular some of its subfields such as evolutionary psychology, have been criticized as a collection of controversial just-so stories, with little bearing on reality. Astrophysics, and astronomy and cosmology in general, are often given much more leeway, even though the underlying scientific hypotheses can be just as weak. One reason for this state of affairs is that while evolutionary biology is a relatively young field, astrophysics and astronomy has been around for a long time, and the practitioners are adept at dealing with the issues mentioned above. In the following, we will discuss how the astrophysical community has managed these inconveniences, and how the same solutions can be applied to evolutionary biology as well. Eventually, our foray will lead us to conclude that much of the blanket criticism of evolutionary biology, and by proxy, evolutionary psychology, as fundamentally untestable mumbo-jumbo is unfounded. Granted, some of the research in these fields may have been dubious, but the problem is not fundamental.
One Earth, One Universe, One Big Problem
The most fundamental of the issues in observation is the fact that we only have one Universe that we know of and can observe. Likewise, we only know of one instance where life has evolved out of inanimate matter. It may be that life was born and died out multiple times and on multiple planets or other bodies of the solar system in the past, but the life on Earth is the only instance that we know of. Hence, it is the only we one we have available for observations.
The problem here is that both the evolution of life and the evolution of the Universe, stars, galaxies and so on are processes, or sequential changes in time of something we can observe. Both processes depend on the initial conditions, that is they start from some specific state of affairs. They also depend on random occurences along the way, with quantum and classical uncertainty guaranteeing that no two Universes or Trees of Life starting from the same initial conditions end up exactly the same. Unless there is no fundamental quantum incertainty, which seems unlikely.
The underlying predicament here is that to understand a process through observations, we would like to be able to observe a whole ensemble of them. If we start some process numerous times with the same initial conditions, we might find that some end results are more common than others. We could then say something about the probability of the outcomes of the process, given some initial conditions. Naturally, we would then repeat the experiment with many other initial conditions, to finally arrive at a better understanding of what the process does in general.
The Life As We Know it was born on Earth (debatable), in very specific conditions, on a very specific planet, which has since suffered from tectonic and volcanic events, as well as impacts by other astronomical bodies. Furthermore the Earth during all this time has been perturbed on its orbit by other celestial bodies, and the Sun has gone through its own changes, both affecting life on Earth. We see that the life on Earth is a unique realization of the process in which life is born and evolves. We may feel then that by observing it, there is little we can say about the birth of life and evolution in general. However, all is not lost.
Cosmologists face a similar kind of fundamental problem. There is only one Universe that we can observe and make deductions about. However, by careful study of all the component processes, that is, fundamental physics, we can at least make educated guesses about what other universes with different initial conditions, such as different fundamental constants, might be like. Turns out that in most cases they would be rather boring. For example, composite particles (nucleons, atoms, molecules) might fail to form, and the universe would be filled with only radiation and fundamental particles. Or the lifetime of the universe might end up so short that nothing interesting, such as life, has time to happen.
So, by understanding the component processes of life, we can get at least somewhere. Careful research and prodigious amount of simulation may yet yield essential and general insights into life and evolution. Not only that, but as far as the birth of life is concerned, several laboratory experiments, such as the famous Miller-Urey experiment, have produced interesting results resembling the building blocks of primordial life using only non-organic ingredients. In addition, isolated island ecosystems have allowed us to observe different realizations of the evolutionary process with rather similar initial conditions. Interestingly, the results indicate that oftentimes evolution tends to converge to similar results, in what is known as the Island rule, and other monikers as well.
Laboratory Astrophysics and a problem of time scales
Such a thing hardly exists. The energies, distances and densities involved in fundamental astrophysics are decidedly unattainable in a lab, since they may require such laboratory equipment as, say, supermassive black holes. These objects, sitting in the centers of distant galaxies, may exceed 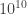

Evolutionary biology is faced with a similar quandary. For one, a replication of the environment affecting the evolution of any macroscopic organism requires a vast amount of real estate, replicated in its complex detail. This environment naturally includes other species as well, which complicates things even further. In addition, the relevant time scale for major evolutionary changes to happen, is apparently much too great for us to wait around to observe. Not to mention that this expected change may not happen at all, since an organism may initially evolve swiftly, but after becoming well-adapted may continue to exist fundamentally unchanged until it is driven extinct (see http://en.wikipedia.org/wiki/Punctuated_equilibrium and related ideas).
Astrophysicists manage their particular set of problems with an indirect observational approach. As an example, we may consider the problem of the evolution of stars. We know that the lifespans of even the most massive stars imaginable are hundreds of thousands of years at the very least. So how, then, can we get an idea of how a star proceeds from its birth to its death? We are saved by the very number of stars we can observe. If we carefully observe thousands of stars, we should statistically expect to see some that are young, some that are old, some in the process of forming and others during their final moments. With our modern telescopes, even the discovery of violent death of a faraway massive star as a supernova is a routine occasion. However, the previous sentence already expressed the problem inherent in this method of observation: we can not be sure that all stars follow a similar evolutionary path. Indeed, it turns out that they don’t, and that their fate is determined by their initial mass and chemical composition. But even this we can discern by the method described, since by collecting a large set of observations, we see that when grouped by some observational characteristics, the stars seem to cluster together in interesting patterns. For example, if we plot an analogue of the observation color of the star and it’s absolute brightness, we end up with the Hertzsprung-Russell diagram (HR-diagram, see Figure 1). In the figure, different evolutionary states of the stars clump together in patterns. The different evolutionary pathways taken by stars of different mass and composition are not immediately obvious, but can be discerned by the aid of theoretical work, and collecting HR-diagrams of different populations of stars. So in this case, we can surmount the problems of time, size and energy scales by observing a great number of objects, and combining the results with theoretical understanding and lab experiments of the component processes involved.

Figure 1. The Hertzsprung-Russell diagram. Source: “HRDiagram” by Richard Powell – The Hertzsprung Russell Diagram. Licensed under CC BY-SA 2.5 via Wikimedia Commons.
In evolutionary biology, we can in principle employ a similar kind of strategy. Since we cannot observe the evolutionary progress of any organism, apart for microscopic ones, we can instead observe a great number of different species, and hope to catch them in different evolutionary states. We can also employ the fossil record, to observe the remains of organisms already extinct. In light of the punctuated equilibrium hypothesis (see above), however, it would require a veritable stroke of luck to catch a species in midst of evolutionary upheaval. We are in luck, though, since we know at least one species where a swift change, at least in evolutionary terms, is occurring: homo sapiens. As an example, the state of human dentition is in flux; the number, age of expression and state of wisdom teeth vary on an individual basis, and even the number of other teeth are affected on non-marginal subsets of people (see hypodontia and hyperdontia for example). In addition, we have the luck of being able to observe different isolated island ecosystems, and study the evolutionary convergences evident in the end results, even if we can’t see the evolutionary process itself happen.
To not rely on luck, we might also try once again to downscale the process, and observe what happens with microscopic organisms like bacteria. We would then like to replicate the success of astrophysics, and use our knowledge of the constituent processes mixed with theoretical understanding, to attain an understanding of the evolutionary process on a grander scale. In principle this is very doable, since a number of different cultures of bacteria can be comfortably fit in a laboratory, and their evolutionary cycle is short enough so that demonstrable changes can be observed within days instead of years. We can then use these observations to get a grasp of the evolutionary processes on a molecular level. However, evolution also happens on the level of a gene, and on the level of a population, in the form of inter-gene, inter-sex and inter-species competition. If evolution happened in a purely statistical manner, driven by environmental pressure and stochastic mutations, it would be easier to understand. Sexual reproduction and the gene-mixing process of meiosis combined with selection and inter-gene competition within both the cellular DNA as well as the mitochondrial DNA make evolution a much more complicated process to understand. A serious complication is that these processes cannot be understood by observing only asexually procreating bacteria, even though they express some behaviors, such as transformation, that mimic the DNA mixing of sexual procreation. Thus in the end, we need to perform a much larger variety of cunning experiments to figure out how all these additional layers of evolutionary mechanisms work together to produce the diversity that we can see now, and observe in the fossil record. A good if slightly controversial book on these “lab experiments” is The Red Queen: Sex and the Evolution of Human Nature.
Conclusion
While the two scholarly disciplines discussed here might seem very unlike, they both grapple with similar kinds of fundamental problems. The issues are fundamental in the sense that they threaten the Popperian falsifiability, on which a sound scientific method must rely on, to a degree. Of course, there are other fields of natural science, such as geology, that face similar restrictions imposed by their objects of study. However, the parallels discussed here are made more striking by the difference of the fields we find them in. More striking are also the fundamental similarities in the means by which scientists try to circumvent these problems. Indeed, we can condense these to an abstract list of things to do:
- break problem down to component processes, and study these by theory and experiment
- substitute observations in time domain with point observations distributed over a large number of (hopefully) related phenomena
- simulate prodigiously
In addition, we have slightly scratched the surface of the scientific underpinnings of evolutionary biology. Far from being a pseudoscience of artful tale-telling, it is a solid field of natural science where well-posed experiments can be done and are being done, and where theories can be falsified and supplanted with other theories. By an extension, evolutionary psychology can then also be built on a basis of solid science. And since it (together with neuropsychology) can potentially teach us a great deal of the human condition and everyday experience, evolutionary psychology will most probably be an often discussed topic here.

{kind=link}